生物界において個体、また個体を形成する器官、組織、細胞などの大きさがいかに決定されるかという問題は基礎、応用研究上非常に重要ですが、その制御機構についてはほとんど解明されていません。私たちはこの問題を特に植物の器官、細胞の大きさ制御の視点から解明することを目指しています。植物の器官や細胞の大きさは外因性、内因性の様々な要素からなる複雑な制御ネットワークによって規定されています。モデル植物シロイヌナズナのloss-of-function、gain-of-function変異体を用いてこれらのネットワークに関与する遺伝子群を同定し、高等植物の大きさを決定する制御システムを解明していきます。またこれらの基礎研究から得られた成果を利用して作物の生産性を向上するための新手法を確立することを目指します。
![]()
細胞分裂周期から核内倍加周期へ
細胞分裂周期から核内倍加周期への移行を制御する分子機構
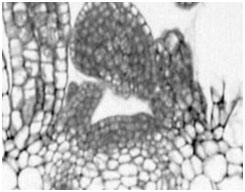 <b>シロイナズナの茎頂メリステム</b><br>茎頂メリステムの細胞は数回分裂を繰り返した後、核内倍加周期に移行し、分化、伸長し始める。 高等植物は地上部の茎の頂点(茎頂)と根の先端(根端)にメリステムと呼ばれる分裂組織を持ち、花や葉、根等の器官の形成や生長に必要な新しい細胞を生産、供給しています。こうしたメリステムでの細胞増殖は、染色体の複製と細胞質分裂が交互に繰り返される細胞分裂周期 (mitotic cell cycle) を経て行われます。多くの植物細胞は数回の細胞分裂を繰り返した後、細胞質分裂を経ないで染色体複製だけが繰り返される核内倍加周期 (endoreduplication cycle / endocycle) へと移行し、細胞内のDNA量 (核相 / ploidy) を増加させ始めます。これまでの研究からメリステムの細胞が核内倍加周期へ移行する時期が、これらの細胞が最終的な分化を開始したり、細胞伸長を開始する時期にしばしば一致することが知られています。このため正常なメリステムを形成、維持するためには、細胞が正しい時期に正しい場所で細胞分裂周期から核内倍加周期へと移行する必要があります。
<b>シロイナズナの茎頂メリステム</b><br>茎頂メリステムの細胞は数回分裂を繰り返した後、核内倍加周期に移行し、分化、伸長し始める。 高等植物は地上部の茎の頂点(茎頂)と根の先端(根端)にメリステムと呼ばれる分裂組織を持ち、花や葉、根等の器官の形成や生長に必要な新しい細胞を生産、供給しています。こうしたメリステムでの細胞増殖は、染色体の複製と細胞質分裂が交互に繰り返される細胞分裂周期 (mitotic cell cycle) を経て行われます。多くの植物細胞は数回の細胞分裂を繰り返した後、細胞質分裂を経ないで染色体複製だけが繰り返される核内倍加周期 (endoreduplication cycle / endocycle) へと移行し、細胞内のDNA量 (核相 / ploidy) を増加させ始めます。これまでの研究からメリステムの細胞が核内倍加周期へ移行する時期が、これらの細胞が最終的な分化を開始したり、細胞伸長を開始する時期にしばしば一致することが知られています。このため正常なメリステムを形成、維持するためには、細胞が正しい時期に正しい場所で細胞分裂周期から核内倍加周期へと移行する必要があります。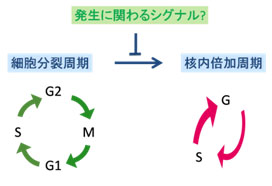 細胞分裂周期から核内倍加周期への移行はどのようなシグナルによって制御されているのだろうか? 私達のグループではシロイナズナを材料として、植物細胞の核内倍加周期への移行がどのように制御されているかを研究しています。またこうした細胞周期制御が細胞の発生、分化過程にどう関与するかを明らかにしようとしています。私達は近年核内倍加周期への移行を抑制する因子の単離を目指したスクリーニング系を構築し、これまでに数系統の新規突然変異体high ploidy (hpy)を単離しました。また最近これらの変異体の解析を通じて、SUMO E3 ligaseをコードするHIGH PLOIDY 2 (HPY2)が核内倍加周期への移行制御及びメリステム維持に必要であることを解明しました。
細胞分裂周期から核内倍加周期への移行はどのようなシグナルによって制御されているのだろうか? 私達のグループではシロイナズナを材料として、植物細胞の核内倍加周期への移行がどのように制御されているかを研究しています。またこうした細胞周期制御が細胞の発生、分化過程にどう関与するかを明らかにしようとしています。私達は近年核内倍加周期への移行を抑制する因子の単離を目指したスクリーニング系を構築し、これまでに数系統の新規突然変異体high ploidy (hpy)を単離しました。また最近これらの変異体の解析を通じて、SUMO E3 ligaseをコードするHIGH PLOIDY 2 (HPY2)が核内倍加周期への移行制御及びメリステム維持に必要であることを解明しました。
植物細胞の生長を制御する分子機構
植物細胞の生長を制御する分子機構
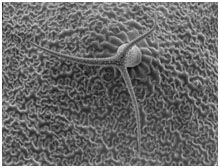 <b>シロイヌナズナのトライコーム</b><br>葉の表皮細胞が分化して形成されるトライコームは、単細胞からなっており、数百倍に巨大化する。 植物細胞が最終的に到達する大きさは、植物種や細胞種によってほぼ一定であるため、植物種固有の遺伝情報によって細胞生長を規定する仕組みが存在することが予想されます。植物細胞の伸長生長は、液胞への吸水と細胞壁の進展による細胞の容積増大によって引き起こされます。また多くの植物細胞の生長には、核内倍加周期によって細胞内のDNA量(核相)が増加することが深く関係しています。これまでに植物細胞の伸長生長を促進する働きを持つ遺伝子は多数単離され、それらの植物体内での機能についての理解は進んできました。しかし植物細胞の生長を積極的に抑制し、停止するような仕組みが存在するかどうかは知られていませんでした。
<b>シロイヌナズナのトライコーム</b><br>葉の表皮細胞が分化して形成されるトライコームは、単細胞からなっており、数百倍に巨大化する。 植物細胞が最終的に到達する大きさは、植物種や細胞種によってほぼ一定であるため、植物種固有の遺伝情報によって細胞生長を規定する仕組みが存在することが予想されます。植物細胞の伸長生長は、液胞への吸水と細胞壁の進展による細胞の容積増大によって引き起こされます。また多くの植物細胞の生長には、核内倍加周期によって細胞内のDNA量(核相)が増加することが深く関係しています。これまでに植物細胞の伸長生長を促進する働きを持つ遺伝子は多数単離され、それらの植物体内での機能についての理解は進んできました。しかし植物細胞の生長を積極的に抑制し、停止するような仕組みが存在するかどうかは知られていませんでした。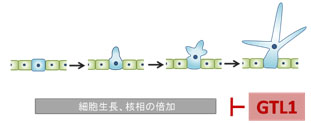 <b>トライコーム形成過程でのGTL1の役割</b><br> 隣り合う表皮細胞列の中から、一部の細胞(青色でマークした細胞)がトライコームに分化し、生長を始める。GTL1遺伝子はトライコーム細胞が最終的な大きさに達する時期に発現し、細胞生長を終了させる。トライコームの細胞生長には核相の倍加現象が深くかかわっており、GTL1は核内倍加の進行にブレーキをかける。 単細胞からなるシロイナズナの葉のトライコームは、表皮細胞から分化後数百倍まで巨大化するため、植物の細胞伸長を研究するモデル系としてよく用いられます。私達はこの系を利用し、細胞が野生型よりもさらに巨大化する突然変異体の探索を試みました。シロイナズナのFOX (Full-length cDNA over-expresser gene hunting system) 変異体コレクション (Ichikawa et al, Plant J 2006) からトライコームが巨大化する突然変異体を単離し解析を行った結果、植物に特有なトライへリックス型転写因子GT-2-LIKE 1 (GTL1)が核相依存的な細胞伸長の抑制に関与することを明らかにしました。
<b>トライコーム形成過程でのGTL1の役割</b><br> 隣り合う表皮細胞列の中から、一部の細胞(青色でマークした細胞)がトライコームに分化し、生長を始める。GTL1遺伝子はトライコーム細胞が最終的な大きさに達する時期に発現し、細胞生長を終了させる。トライコームの細胞生長には核相の倍加現象が深くかかわっており、GTL1は核内倍加の進行にブレーキをかける。 単細胞からなるシロイナズナの葉のトライコームは、表皮細胞から分化後数百倍まで巨大化するため、植物の細胞伸長を研究するモデル系としてよく用いられます。私達はこの系を利用し、細胞が野生型よりもさらに巨大化する突然変異体の探索を試みました。シロイナズナのFOX (Full-length cDNA over-expresser gene hunting system) 変異体コレクション (Ichikawa et al, Plant J 2006) からトライコームが巨大化する突然変異体を単離し解析を行った結果、植物に特有なトライへリックス型転写因子GT-2-LIKE 1 (GTL1)が核相依存的な細胞伸長の抑制に関与することを明らかにしました。
器官サイズ決定に関わる分子機構
植物の器官サイズを決定する分子機構
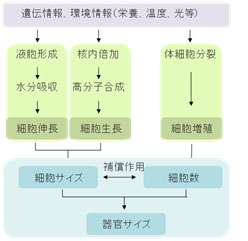 植物の器官サイズは構成細胞のサイズと数のバランスによって規定される。 “器官サイズがいかにして決まるか”という問題は、基礎科学においても応用研究においても非常に重要です。ある器官の最終的な大きさは、それを構成する細胞の分裂頻度や伸長程度によって規定されます。最近の研究から、これらの過程を制御する遺伝子が同定され始めていますが、これらの個別の因子がどういった機能を持ち、どのように器官サイズを規定するのかについての統合的な理解はまだ進んでいません。
植物の器官サイズは構成細胞のサイズと数のバランスによって規定される。 “器官サイズがいかにして決まるか”という問題は、基礎科学においても応用研究においても非常に重要です。ある器官の最終的な大きさは、それを構成する細胞の分裂頻度や伸長程度によって規定されます。最近の研究から、これらの過程を制御する遺伝子が同定され始めていますが、これらの個別の因子がどういった機能を持ち、どのように器官サイズを規定するのかについての統合的な理解はまだ進んでいません。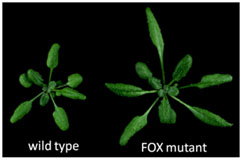 野生型の植物体(左図)よりも大きな葉を持つFOX変異体の一例。 植物における器官サイズ制御機構の解明を進めるために、私達はシロイナズナのFOX (Full-length cDNA over-expresser gene hunting system) 変異体コレクション (Ichikawa et al, Plant J 2006) から、野生型に較べて器官サイズが大きくなったり、小さくなったりする突然変異体を単離しました。現在これらの原因遺伝子の機能解析を進め、器官サイズを決定する新規な制御過程を明らかにしようとしています。
野生型の植物体(左図)よりも大きな葉を持つFOX変異体の一例。 植物における器官サイズ制御機構の解明を進めるために、私達はシロイナズナのFOX (Full-length cDNA over-expresser gene hunting system) 変異体コレクション (Ichikawa et al, Plant J 2006) から、野生型に較べて器官サイズが大きくなったり、小さくなったりする突然変異体を単離しました。現在これらの原因遺伝子の機能解析を進め、器官サイズを決定する新規な制御過程を明らかにしようとしています。
記事
 RIKEN Research:植物のサイズアップ
RIKEN Research:植物のサイズアップ 理研ニュース:植物の大きさはどう決まる?
理研ニュース:植物の大きさはどう決まる?
プレスリリース
 植物細胞の脱分化を促進するスイッチ因子を発見(2011年3月)
植物細胞の脱分化を促進するスイッチ因子を発見(2011年3月) 植物細胞の大きさを調節する新たな遺伝子「GTL1」を発見(2009年9月)
植物細胞の大きさを調節する新たな遺伝子「GTL1」を発見(2009年9月) 細胞分裂の調節に必須の新しい「HPY2」遺伝子を発見(2009年8月)
細胞分裂の調節に必須の新しい「HPY2」遺伝子を発見(2009年8月) DNAの量によって植物の大きさが決まる新たな仕組みを解明(2007年12月)
DNAの量によって植物の大きさが決まる新たな仕組みを解明(2007年12月)













